research
Overview
I use multy electrode array electrophysiology on awake and listening animals to record the responses of their auditory cortex to sounds. I then use computational methods to model how past and ongoing sounds are represented by neurons in a population and putative circuit of neurons. With this I hope to understand how the brain accumulates, transforms and relays information, representations and abstractions, and how we can draw inspirations from these algorithms to engineer smarter artificial intelligence.
Projects
Contextual representation and sparse coding
Keeping track of both past and ongoing stimuli is necessary for computation and cognition. However, how this working memory is represented by the auditory cortext, and how this context of past stimuli modulates current response remains unknown. We have found long-lasting contextual modulation in both the primary auditory cortex (A1) and the peri ectosylvian gyrus (PEG), a secondary auditory cortex region. This contextual modulation lasts hundreds of milliseconds, past beyond the archetypal integration window of A1 and PEG spectro-temporal receptive fields (STRF). Furthermore, it is distributed in a sparse code across neurons in a local population. Finally, we have shown that models containing local synaptic connections, surrogate for recurrent connections, better capture the response of neurons to sound, alongside the aforementioned contextual modulation. See A sparse code for natural sound context in auditory cortex .
Short term plasticity and stimulus specific adaptations
There is extensive work on the physiology behind the multiple mechanism -both synaptic and neuronal- underlying short term plasticity (STP), the ability of a neuron to modulate its response as a consequence of prior activity. However, we don't fully understand how this STP acts in the context of populations and circuits of neurons encoding auditory stimuli. We have shown how short term plasticity happening independently for different spectral input channels can better predict the recorded neural activity, and is also capable of recapitulating stimulus specific adaptation, a response property seen across brain regions and species. See Spectral tuning of adaptation supports coding of sensory context in auditory cortex
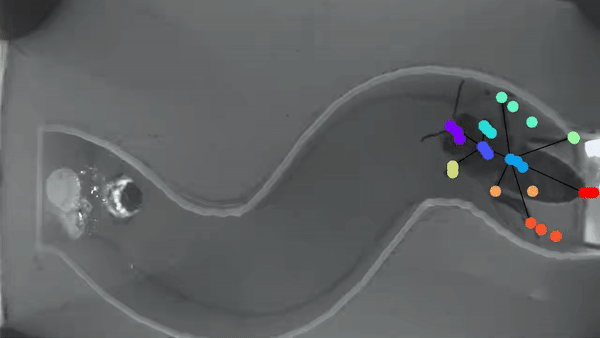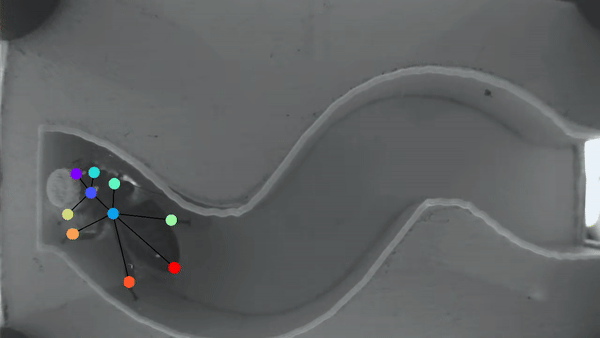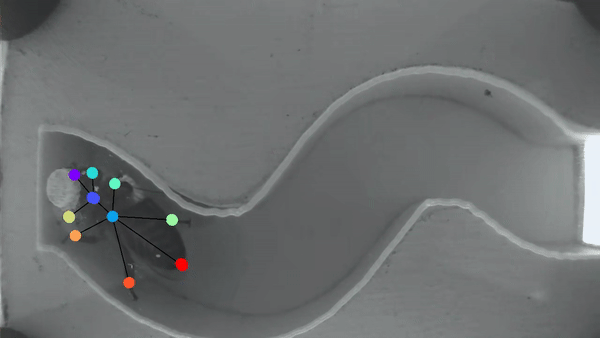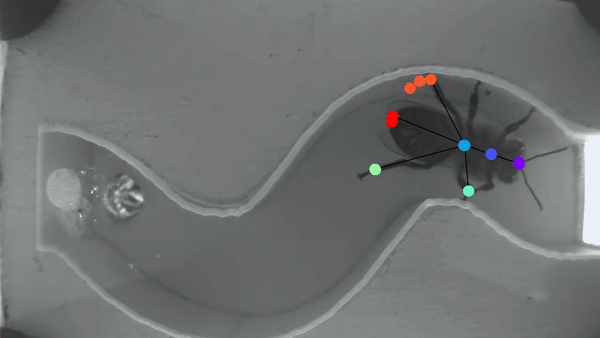
Bee manipulation of flowers constrains foraging preference
A bee prefers visiting flowers of the same type. This “Flower constancy” is advantageous for the plant, however the advantages for the bee are less clear. A classic explanation lies in the cognitive cost of learning to discriminate and remember the reward of multiple flower types. However less is known about the cognitive cost of learning different manipulations required to gain access to flower nutrients. In order to understand this manipulation cost, we made a detailed characterization of flower harvesting behaviors. Videos of bees mid-harvest were analyzed using available deep learning tools to generate a dictionary of constitutive movements, that are assembled into distinct flower manipulations. We predict that flowers with very different manipulation will necessitate an increased cognitive load and thus generate flower constancy.